Bone is living, dynamic tissue that affects three main functions of the body: skeletal homeostasis (providing structure to house the internal organs as well as a system of pulleys and levers to move the body), mineral homeostasis (storing and releasing ions), and hematopoiesis (accommodating the machinery of blood cell formation). As living tissue, bone is in a state of continuous flux and renewal; osteoclastsresorb bone, and osteoblasts create new bone. This process allows ions to be released and stored. It also repairs areas of structural damage.
Most bone forms according to the cartilage model, a process called endochondral ossification. The process of bone growth and development, therefore, is one of transformation from a cartilage skeleton in utero to one made entirely of bone in the adult. The pediatric skeleton contains remnants of this cartilage skeleton, the cartilaginous growth plate or physis. The physis is active metabolically and allows the pediatric skeleton to grow longitudinally; however, it is also structurally weaker than bone, which makes the pediatric skeleton susceptible to a class of diseases and injuries not seen in the adult.
Bone Formation
Bone forms by one of two mechanisms: endochondral or intramembranous ossification. Endochondral ossification is the most common form, producing the long bones; “long” refers to the longitudinal orientation of the bone. (Even the distal phalanx of the fifth toe is a long bone.) The flat bones, such as the skull and clavicle, form via intramembranous ossification. The key distinction between the two processes is that endochondral ossification employs a cartilage template whereas intramembranous ossification does not.
Morphogenesis, or the development of the skeleton in utero, begins in the fourth week of development, at which time limb buds form in the embryo. When the fetus reaches the sixth week of development, mesenchymal cells (cells from the middle of the three germ cell layers) appear within the central region of the limb buds. These cells then differentiate into cartilage-forming chondroblasts, which elaborate a cartilage template of the bony skeleton. After these cells die, blood vessels migrate into the mass of matrix, bringing the osteoblasts, or bone-forming cells. Osteoblasts synthesize and secrete a collagen-based material called osteoid; this organic bone matrix then becomes calcified.
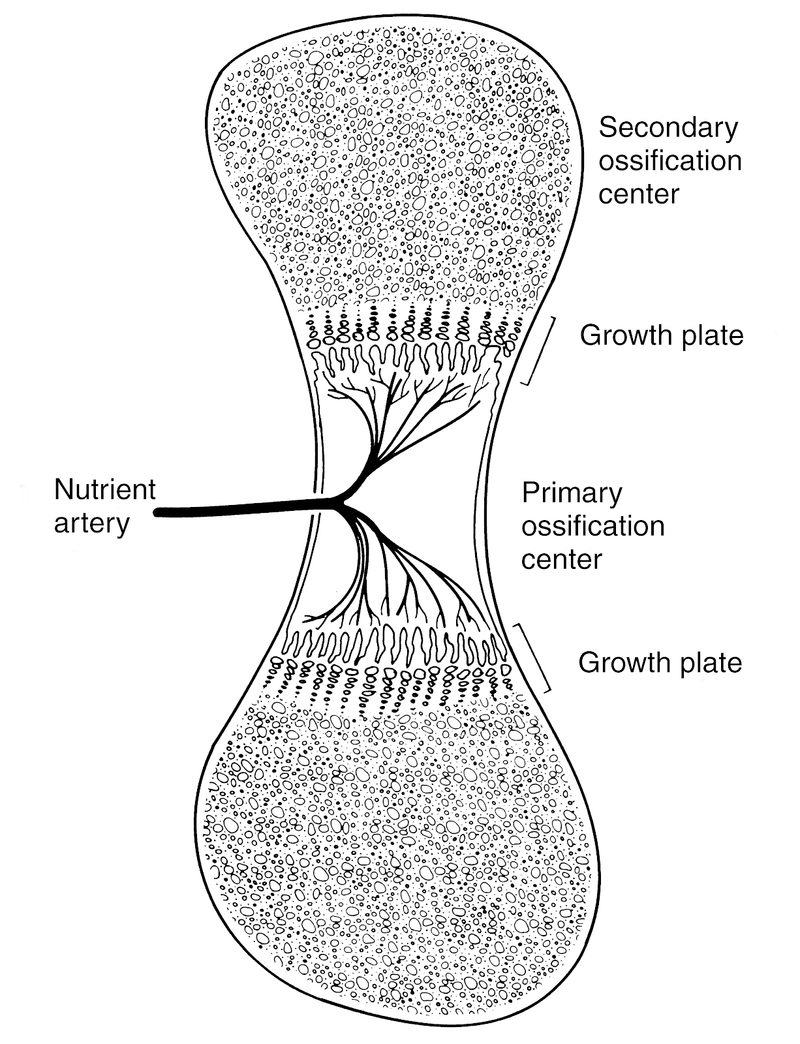
A region of calcified cartilage called the primary center of ossification forms midway between the two ends of the long bone. Calcification begins here and continues longitudinally toward the ends of the long bone, a process called interstitial growth, which takes place entirely within the cartilage model.
As interstitial growth continues, the primary ossification center enlarges, migrating toward but never ultimately reaching the ends of the bone. At the edge of the advancing primary ossification center, a specialized structure, the physis, or growth plate, forms (Fig. 1).

Figure 1 The primary ossification center stands at the midportion of the developing bone. Toward each end lies a growth plate (physis) and beyond it, a secondary ossification center will develop.
(Reproduced with permission from Brighton CT: The growth plate. Orthop Clin North Am 1984;15:571.)
The physis remains cartilaginous throughout adolescence to allow longitudinal growth of the maturing skeleton.
Just beyond the physeal growth plate, in the regions of the long bones farthest away from the primary ossification center, lies the chondroepiphysis. This area will become a secondary center of ossification. Some bones, such as the proximal and distal humeri, have several secondary centers of ossification that appear in a sequential manner during childhood and then fuse just before skeletal maturity to yield the adult form of the bone.
The primary and secondary ossification centers on either side of the physis define the following regions of the formed bone: the diaphysis (shaft), representing the primary center; the metaphysis, the flared region just below the physis; and the epiphysis, which is located across the physis just below the articular cartilage lining the joint (Fig. 2).

Figure 2 Schematic representation of the regions of the long bones in the tibia.
Within the diaphysis, a central core called the medullary canal is hollowed out by resorption, creating a cavity for the future home of the blood-forming cells.
As interstitial growth lengthens the bone, a parallel process of appositional growth widens the bone. Appositional growth takes place under the periosteum, a sleeve of connective tissue surrounding the shaft of the bone. The periosteum is a thick and distinct membrane in children but remains present in an attenuated form throughout life. It is the source of cells used for fracture healing and the source of some new bone formation.
Endochondral ossification continues long after the skeleton ceases longitudinal growth. Instead of stopping the healing process at the scar formation stage (as most tissues are apt to do), the repair of bone continues via endochondral ossification, which converts cartilaginous scar into new bone. Thus, it is quite possible that many years after a long bone fracture, no structural evidence of the original injury exists.
Growth Plate Anatomy and Function
At the physis, chondrocytes proliferate, mature, and secrete extracellular matrix, which eventually ossifies. The physis is polar: moving away from the epiphysis and joint surface toward the metaphysis, there is increasing maturation of the cells. Just below the epiphysis is a zone of immature resting cells; just above the metaphysis, calcification takes place in the setting of programmed cell death (apoptosis) of the hypertrophic chondrocytes. New bone formation thus occurs at the point farthest from the articular surface, at the junction between the physis and the metaphysis. This region is named the zone of provisional calcification.
The physis itself is divided into three major zones: the reserve zone, the proliferative zone, and the hypertrophic zone (Fig. 3).

Figure 3 The zones of the growth plate. Note that the epiphysis is above the reserve zone and the metaphysis is below the hypertrophic zone.
(Reproduced with permission from Brighton CT: Structure and function of the growth plate. ClinOrthop 1978;136:24.)
Each one of these zones has a distinct histology directly related to its function. The zones are ordered such that metabolic activity and maturation increases the farther the cells are located from the joint surface.
Closest to the surface (ie, nearest the epiphysis) is the reserve zone. The cells in this zone produce cartilaginous matrix, primarily in the form of type II collagen, which is used for eventual ossification into bone. These cells do not actively divide, nor are they very metabolically active; accordingly, they have the poorest blood supply.
Below the reserve zone, moving toward the metaphysis, is the proliferative zone. Within this zone, the cells are stacked in columns. These columns of cells synthesize proteoglycans, thereby contributing to the extracellular matrix. This region, characterized by synthesis and cell division, has the most extensive blood supply within the growth plate. This blood supply provides nutrition, of course, but also allows hormonal signals to reach their targets in the growth plate effectively.
The third zone of the growth plate, the hypertrophic zone, lies closest to the calcified bone of the metaphysis. The cells in this zone are unusually large and plump; hence, the name “hypertrophic.” This zone of the growth plate is highly active metabolically, even though it has a poor blood supply. It therefore relies on anaerobic metabolism and uses stored glycogen as its source of energy. It is also the region that participates in mineralization of the cartilage. Calcium is stored in the cells in the upper levels of the hypertrophic zones. In the lower levels of the hypertrophic zone, these cells liberate their calcium in order to assist in matrix mineralization.1
The actual process of matrix mineralization occurs at the interface between the hypertrophic zone and the metaphysis: the zone of provisional calcification. Within this area, vascular invasion allows osteoblasts to arrive and replace the calcified cartilage with bone. This bone is a primitive, less-organized form called woven bone. In time, this tissue will be replaced by mature lamellar bone via the process of bone remodeling. The distinction between woven and lamellar bone is one of material orientation: the fibers of woven bone are haphazard, whereas lamellar bone aligns the structure in the direction of load.
Blood does not flow easily through the physis; the intramedullary blood supply does not reach the epiphysis or secondary centers of ossification. Accordingly, in children, whose growth plates are open, the main blood supply to the epiphysis is a direct epiphyseal artery. This artery loses its prominence once the growth plates close at skeletal maturity.
Numerous circulating hormones affect growth plate activity. Thyroid hormone (thyroxine), growth hormone, parathyroid hormone (PTH), calcitonin, and testosterone are among the hormones used to regulate growth plate physiology. These hormones can stimulate matrix synthesis, cell division, and calcification (and thus growth plate closure). The precise method and zone of action of these hormones are beyond the scope of this text. Even without entirely understanding their mechanisms of action, however, it is clear that abnormalities of these hormones can significantly affect the development of the human skeleton.2
Bone Cells
Three types of cells, osteoblasts, osteocytes, and osteoclasts, are essential to the development of bone and its dynamic remodeling (ie, the process of bone resorption and deposition that releases and deposits ions, adapts the bone to new loads, and repairs microscopic damage). All three types of cells can be seen in any given area of bone, and many metabolic bone diseases are caused by loss of regulation, resulting in disorders of bone formation or remodeling.
Osteoblasts
Osteoblasts are bone-forming cells. Although this is a useful definition for the purpose of discussion, osteoblasts do not actually form complete bone. Rather, they synthesize the organic (ie, nonmineral) component of bone, the primary element of which is osteoid (a protein matrix of type I collagen). Osteocalcin and bone sialoproteins are other organic products of osteoblasts. Through these products, the osteoblasts regulate the mineralization of osteoid. Bone, therefore, is a composite of organic products of cellular synthesis and mineral deposited by extracellular matrix calcification.3,4
Because osteoblasts are metabolically active, they typically contain a large Golgi apparatus (Fig. 4).

Figure 4 Light (A) and electron (B) microscope views of the osteoblast.
(Reproduced from Bostrom MPG, Boskey A, Kaufman JK, Einhorn TA: Form and function of bone, in Buckwalter JA, Einhorn TA, Simon SR (eds): Orthopaedic Basic Science: Biology and Biomechanics of the Musculoskeletal System, ed 2. Rosemont, IL, American Academy of Orthopaedic Surgeons, 2000, p 323.)
This organelle is responsible for secreting type I collagen, the prime component of the bone matrix. Osteoblasts also contain a large nucleus that is usually oriented away from the osteoid-producing side of the cell. In addition to collagen, osteoblasts also produce an enzyme called alkaline phosphatase. The precise function of alkaline phosphatase is still unknown; however, based on the observation that patients who are producing bone (ie, those with fractures or bone-forming tumors) have increased serum levels of alkaline phosphatase, researchers reasonably conclude that it plays some role in the mineralization of osteoid. Alkaline phosphatase is therefore a useful clinical marker of bone formation activity.
Osteoblasts are derived from the mesenchymal cell line. Once a mesenchymal cell becomes committed to the bone cell lineage, it is known as a preosteoblast. These preosteoblasts become osteoblasts when properly stimulated. The precise signals responsible are not known, but substances shown to have a role in the regulation of the differentiation of this cell line include bone morphogenetic proteins, growth factors, interleukins, insulin-derived growth factor, and platelet-derived growth factor.
The organic matrix secreted by osteoblasts is primarily type I collagen. Approximately 10% of the mass of the osteoid is composed of other matrix proteins and growth factors. Mineralization of the matrix occurs via a controlled process that has not been completely identified. It is known, however, that calcium must be locally concentrated (to form precipitates) and that energy is required to initiate the process. Some of the noncollagenous matrix proteins apparently promote or inhibit mineralization and modulate crystal size.
Osteocytes
Osteocytes, the cells of established bone, are derived from osteoblasts. In fact, an osteocyte is simply an osteoblast that has surrounded itself with bone matrix. Of course, the morphology of the cell changes: because the osteocyte is less metabolically active, it contains fewer organelles, and the nucleus occupies a greater proportion of the entire cell body. Although osteocytes are, by definition, surrounded by osteoid, they are able to communicate with other distant osteocytes through long cell processes traveling through the canaliculi of bone. This network of osteocytes can regulate its metabolic environment and respond to signals applied to the surface of the bone.
Osteoclasts
Whereas osteoblasts and osteocytes are responsible for the production of bone, osteoclasts break it down. Osteoclasts are large cells, derived from stem cells in the bone marrow that give rise to the monocyte-macrophage cell line. Macrophages and osteoclasts, both formed by the fusion of monocyte precursors, are multinucleated. When examined under light microscopy, osteoclasts are distinguished easily from other bone cells by their nuclear morphology (Fig. 5)

Figure 5 Light photomicrograph of a multinucleated osteoclast with a resorption (Howship’s) lacuna.
(Reproduced from Bostrom MPG, Boskey A, Kaufman JK, Einhorn TA: Form and function of bone, in Buckwalter JA, Einhorn TA, Simon SR (eds): Orthopaedic Basic Science: Biology and Biomechanics of the Musculoskeletal System, ed 2. Rosemont, IL, American Academy of Orthopaedic Surgeons, 2000, p 324.)
and ruffled borders. This ruffled border increases the surface area of the cell in contact with bone, and its presence indicates an “active” osteoclast.
Osteoclasts reside in pits on the surface of bone called Howship’s lacunae. The stimulus for osteoclasts to begin bone resorption is provided by osteoblasts. In response to signals for bone resorption (such as PTH), osteoblasts secrete neutral proteases that expose the surface of the bone. Osteoclasts then work on this surface by attaching to the bone by means of receptors known as integrins. Their ruffled borders then secrete proteolytic enzymes and hydrogen ions (acidifying the local environment) that degrade the bone matrix. In this acidic environment, calcium hydroxyapatite crystals, which make up the mineral content of bone, become more soluble and are released into the circulation.
Normal Histology and Composition
Bone is efficient. While forming and remodeling, mass is kept to a minimum. Bone is divided into two structural types: cortical (bark-like) and trabecular. Cortical bone is dense, with its collagen aligned in the direction of applied forces; the cortex of bone is found at the periphery and is responsible for skeletal homeostasis. Trabecular bone is characterized by lattices of bone spicules, which are also aligned in the direction of load but with considerably more empty space than cortical bone. This type of bone, therefore, has a large surface-to-volume ratio and as such is best able to answer the demands of mineral homeostasis (as noted above, osteoclasts, which release mineral ions, require a surface on which to operate). Thus, trabecular bone is the area most likely affected by metabolic bone diseases. Trabecular bone also contributes to efficient mechanical support by orienting its spicules in the direction of load (Fig. 6).

Figure 6 Trabecular bone is characterized by a three-dimensional lattice of bony spicules with considerable space among them.
(Reproduced with permission from Gibson JL: The mechanical behavior of cancellous bone. J Biomech 1985;18:317-328.)
Cortical bone, although dense, has its own microanatomy. The basic unit of cortical bone is the osteon. An osteon is a central canal housing a vascular channel surrounded by rings of lamellar bone (Fig. 7).

Figure 7 A bone osteon has a central core surrounded by concentric rings of lamellar bone.
(Reproduced from Bostrom MPG, Boskey A, Kaufman JK, Einhorn TA: Form and function of bone, in Buckwalter JA, Einhorn TA, Simon SR (eds): Orthopaedic Basic Science: Biology and Biomechanics of the Musculoskeletal System, ed 2. Rosemont, IL, American Academy of Orthopaedic Surgeons, 2000, p 321.)
Osteons communicate with each other via side-to-side canals. Osteons allow even cortical bone to remodel by maintaining the tissue’s blood supply; they are likely to play a mechanical role as well, such as stopping the propagation of cracks within the cortex.
Skeletal Homeostasis
The resorption and removal of bone by osteoclasts shapes bones during development and continues throughout life. In the healthy state, bone resorption is coupled with new bone deposition, and the paired process is called bone remodeling. Bone remodeling in the adult frees mineral from the skeleton and removes microscopic areas of damaged bone. Thus, bone remodeling contributes to both mineral homeostasis and skeletal homeostasis, although mineral homeostasis takes precedence: insufficient amounts of calcium can cause cardiac arrhythmias and sudden death, whereas the transient loss of bone structure, however important, does not typically cause rapid and devastating consequences.
Bone remodeling is initiated by both chemical and mechanical signals. The primary chemical signal is PTH, which is mobilized in response to low serum calcium. Estrogen, too, is an important chemical signal. It helps ensure that the actions of osteoclasts are coupled with those of osteoblasts. When an estrogen shortage occurs, the activities of osteoclasts may exceed those of osteoblasts, resulting in decreased skeletal mass over time. This is the phenomenon that underlies perimenopausal osteoporosis. Testosterone also affects bone metabolism via osteoblast function, but its role is less clear than the role of estrogen.
Skeletal signals in the form of physical stresses can also stimulate bone remodeling. Bone is deposited in areas where load bearing is high, a process known as Wolff’s law. Bone deposition may be mediated by direct stress receptors on the bone cells or by the electrical potentials created by deforming (ie, bending) the bone under load bearing. The converse of Wolff’s law also applies to bone remodeling: bone will be resorbed in areas not subject to stress. Accordingly, disuse will lead to significant bone loss. This has practical relevance. For example, a bone that is casted for treatment of a fracture will be weaker than normal, even in the areas that were not fractured.
Bone remodeling requires biologic substrate (material) and energy (power). Thus, bone remodeling takes place only in areas with an adequate blood supply. If an area of bone infarcts (ie, is cut off from circulation and dies), then that area will not remodel. This occurs in osteonecrosis, a process in which the inability to repair subchondral bone eventually leads to fracture and collapse within the joint.
Hematopoiesis
Hematopoiesis, the process of forming blood cells, normally occurs in tandem with the processes of skeletal and mineral homeostasis and not in opposition to them. Nonetheless, diseases of hematopoiesis may impact bone health. Specifically, hematologic malignancies are often found in bone. Also, because the bone is part of the circulatory system (albeit a place where circulation moves sluggishly), metastatic tumor deposits are often found in bone as well. Likewise, circulating bacteria may lodge in the bone causing osteomyelitis.
Mineral Homeostasis
Calcium
Calcium is a key mineral that contributes to the structural strength of the bone, but it has a myriad of other functions in the body as well, including participating in enzyme activation, nerve transmission, and muscle contraction.5 In addition, it functions as an intracellular messenger and as a cofactor in the coagulation cascade. Accordingly, the precise concentration of active (ionized) calcium must be tightly regulated.
Nearly all (99%) of the 1,000 g of calcium held in the body is stored within the bone in the form of a crystalline calcium phosphate called hydroxyapatite (Ca10 (PO4)6 (OH)2). Apatite crystals are 5 × 80 nm or smaller. In bone, there may be impurities of the crystal, in which other ions, such as potassium, sodium, or magnesium, replace the calcium; these substitutions can affect material properties of the bone.
Only 1% of the body’s calcium is found outside the bone, primarily in the body fluids. Within cells, the concentration of ionized calcium is very low, 0.1% that of the extracellular fluid, as free calcium could form precipitates and kill the cell. In the extracellular fluid, approximately 50% of the calcium is protein bound (ie, attached primarily to albumin) or complexed to negatively charged ions, such as citrate. The other 50% is ionized. It is this ionized fraction that is biologically active.
Calcium enters the body through the intestines and is reabsorbed through the renal tubule. Both processes control the level of circulating calcium ions. However, because dissolution of bone matrix can free calcium for other physiologic needs, much of the body’s minute-to-minute calcium requirements are supplied via bone resorption. Nonetheless, the intestinal absorption of dietary calcium is needed to replenish skeletal stores.
In a typical 24-hour period, 500 mg of calcium are released by the bone via osteoclast resorption, and 500 mg are deposited back into the bone via osteoblast-mediated bone formation. In those 24 hours, approximately 10,000 mg of calcium are delivered to the kidneys, and nearly all of it (98%) is retained, with only 200 mg excreted in the urine. This 200-mg loss must be replaced by calcium in the diet; however, 1,000 mg must be ingested—not 200 mg—because intestinal absorption is imperfect and 800 mg are excreted in the stool. Thus, at the point of balance, 1,000 mg of calcium are ingested and 1,000 mg are excreted. The exchange between the bone and the plasma is similarly balanced (Fig. 8).

Figure 8 Calcium balance.
(Adapted with permission from Bullough PG: Atlas of Orthopedic Pathology. Hampshire, England, Gower Press, 1992, p 7.5.)
Phosphorus
Like calcium, phosphorus is stored in the body primarily in the bone, although nearly 100 g are present in the extracellular fluid, approximately 10 times the amount of extraskeletal calcium. In the body, phosphorus is found mainly in the form of phosphate ions, which play a key role in energy generation. The phosphorylation of adenosine is the primary energy exchange system in the cell. Phosphorous molecules, in the form of phospholipids, are plentiful in cell membranes as well.
The average daily intake of phosphorus is approximately 800 to 1,000 mg in the typical American diet, 60% of which is absorbed in the intestine. This rate of absorption can be increased to 90% via the action of vitamin D, if necessary; urinary excretion also helps maintain phosphorus balance. Bone remodeling can also modulate the serum concentration of phosphorus.
Regulation of Calcium Flux
The three points of calcium flux, the intestine, the kidneys, and the bone, are all regulated and can modify their normal exchange rates in response to metabolic demands. Should a drop in the concentration of serum calcium occur—or, more specifically, a drop in the concentration of biologically active, ionized calcium—the kidneys can retain a greater percentage of the serum calcium they receive for filtration. Similarly, bone can also assist in regulating serum calcium by mobilizing skeletal calcium through the resorption of mineralized bone matrix.
Mediators of Mineral Balance
The primary circulating factors that affect calcium balance are PTH, vitamin D, calcitonin, and calcium itself (Fig. 8).
Parathyroid hormone
PTH is synthesized by the parathyroid glands and stored within the glands themselves. In response to low concentrations of serum calcium, PTH is released, and the synthesis of additional supplies is initiated. PTH exerts its effects on calcium and phosphate metabolism in the bones and the kidneys. In bone, PTH stimulates the release of calcium and phosphate in response to low serum calcium levels. Paradoxically, it is not the bone-resorbing osteoclasts but the bone-forming osteoblasts that have receptors for PTH. Osteoblasts, in turn, signal osteoclasts (via means not completely understood) to increase bone resorption and mobilize calcium and phosphate stores. In the kidneys, PTH stimulates the resorption of calcium, decreases phosphate resorption, and promotes the hydroxylation of inactive vitamin D into its biologically active form.
Vitamin D
Vitamin D is a steroid hormone; as such, it binds to the nucleus of a cell and modifies gene expression and protein synthesis. The function of vitamin D in mineral metabolism is to increase the serum concentration of cal-cium. The body obtains vitamin D directly from diet or through primary synthesis (Fig. 9).

Figure 9 Vitamin D synthesis. Note that both the liver and the kidney hydroxylate inactive forms of cholecalciferol; the kidney’s hydroxylation is under PTH control.
(Reproduced from Bostrom MPG, Boskey A, Kaufman JK, Einhorn TA: Form and function of bone, in Buckwalter JA, Einhorn TA, Simon SR (eds): Orthopaedic Basic Science: Biology and Biomechanics of the Musculoskeletal System, ed 2. Rosemont, IL, American Academy of Orthopaedic Surgeons, 2000, p 354.)
The endogenic synthesis of vitamin D begins in the skin when the skin is exposed to sunlight. Sunlight modifies 7-dehydrocholesterol to form the base (inactive) compound of vitamin D3 (cholecalciferol), which is then transported to the liver where it is hydroxylated. This hydroxylation occurs at what is referred to in chemistry nomenclature as position 25; thus, the molecule at this point in the process is called 25-OH cholecalciferol.
25-OH cholecalciferol is then transported to the kidneys, where it is hydroxylated again. Hydroxylation can occur at either position 1 or position 24; the former is the active form, the latter inactive. The “decision” regarding which position will be hydroxylated depends on the availability of circulating vitamin D and PTH. Low levels of vitamin D or high levels of PTH stimulate hydroxylation at position 1, yielding the active form of the vitamin, 1,25 OH2 vitamin D. Likewise, high levels of vitamin D create a feedback loop of sorts and increase the formation of inactive 24,25 (OH)2 vitamin D.
Bone, the intestine, and the kidneys are the target end organs of vitamin D. Like all steroid hormones, vitamin D arrives at these target organs via the bloodstream. Each organ has specific receptors for vitamin D.6 In the bone, vitamin D stimulates the release of calcium and phosphate. In the small intestine, vitamin D promotes the synthesis of proteins that transport dietary calcium into the circulation. In the kidney, it increases calcium resorption.
Calcitonin
Calcitonin is a peptide that is produced and secreted by the chief cells of the thyroid gland. This peptide has a direct effect on osteoclasts by decreasing their size and inhibiting their ability to resorb bone. Calcitonin is directly regulated by serum calcium level. When serum calcium levels are high, the chief cells respond by producing and secreting more calcitonin in order to decrease bone breakdown and thereby decrease the mobilization of calcium. Calcitonin has been used to treat several diseases in which osteoclast resorption of bone is uncontrolled. It also may have a role in decreasing the long-term effects of osteoporosis.
Research and New Directions
Skeletal Formation
Current investigations attempt to identify the genetic, biochemical, and biomechanical signals that cause the skeleton to be formed, take the shape it does, and stop the formation process when complete. These basic biologic studies add to our knowledge, of course, but they may also suggest why diseases of bone loss or dysfunction occur.
Fracture Healing
Fracture healing is known to recapitulate some of the steps of normal bone formation. Studies of the cellular machinery of bone formation are ongoing and may reveal how fracture healing can be accelerated or enhanced.
Regulation of Bone Metabolism
Osteoporosis is a disease in which bone formation is not coupled perfectly with bone resorption. Ongoing laboratory studies, such as those involving menopause and estrogen loss, attempt to further regulate this process to ensure that bone is not lost from the skeleton even as some natural mediators are altered.
Key Terms
Alkaline phosphatase An enzyme produced by osteoblasts that is believed to play a role in the mineralization of bone
Apoptosis Programmed cell death
Bone remodeling A process that couples bone resorption by osteoclasts with deposition by osteoblasts (new bone cells)
Chondroblasts The cells that form cartilage
Cortical bone Dense bone, literally “bark,” that is responsible for skeletal homeostasis
Diaphysis The shaft of a long bone
Endochondral ossification The formation of bone within a cartilage model
Epiphysis The region of bone across the physis, just below the articular cartilage
Hematopoiesis The process of forming blood cells
Hypertrophic zone A layer of large, plump cells in the physis that assists in mineralization of the cartilage
Infarct An area of tissue that is cut off from its blood supply, becomes ischemic, and dies
Lamellar bone Mature, well-organized form of cortical bone
Medullary canal The relatively hollow central core of a long bone that houses blood-forming cells
Metaphysis The region of bone just below the physis
Mineral homeostasis The function of bone responsible for the storing and releasing of ions, principally calcium
Morphogenesis The development of the skeleton in utero
Osteoblasts The cells that synthesize the organic component of bone; also thought of as the bone-forming cells
Osteoclasts Large cells that resorb bone matrix when activated
Osteocytes The cells of established bone
Periosteum A sleeve of connective tissue that surrounds the shaft of the bone and contributes to fracture healing
Physis The growth plate
Proliferative zone A layer of stacked cells in the physis that synthesizes proteoglycans for the cartilaginous matrix; also has the most extensive blood supply within the physis
Reserve zone A layer of cells in the physis that produces the cartilaginous matrix for ossification into bone
Skeletal homeostasis The function of bone that supplies structural support and movement for the body
Trabecular bone Bone characterized by its “lattice” appearance and is responsible for mineral homeostasis
Wolff’s law A law that states that the growth and remodeling of bone is influenced and modulated by mechanical stresses
Woven bone Primitive, less-organized form of cortical bone
References
1. Zaleske DJ: Cartilage and bone development. Instr Course Lect 1998;47:461-468.
2. Iannotti JP: Growth plate physiology and pathology. Orthop Clin North Am 1990;21:1-17.
3. Tonna EA, Cronkite EP: The periosteum: Autoradiographic studies on cellular proliferation and transformation utilizing tritiated thymidine. Clin Orthop 1963;30:218-232.
4. Baron R, Ravesloot JH, Neff L, et al: Cellular and molecular biology of the osteoclast, in Noda M (ed): Cellular and Molecular Biology of Bone. San Diego, CA, Academic Press, 1993, pp 445-495.
5. Boden SD, Kaplan FS: Calcium homeostasis. Orthop Clin North Am 1990;21:31-42.
6. Walters MR: Newly identified actions of the vitamin D endocrine system. Endocr Rev 1992;13:719-764.